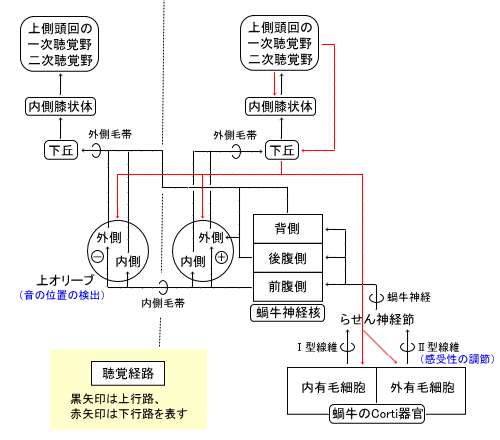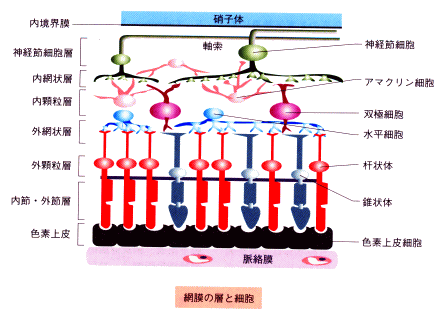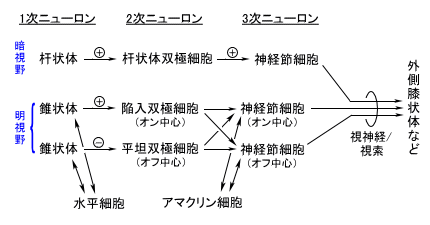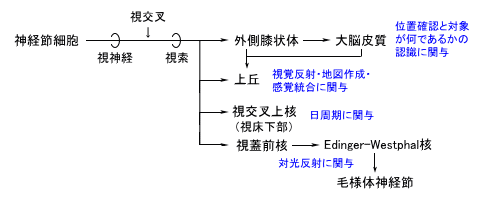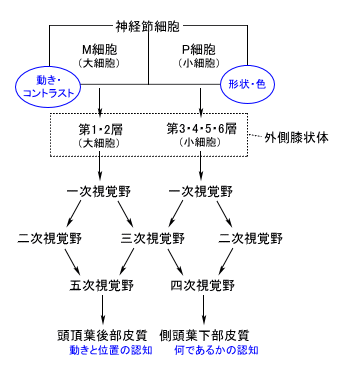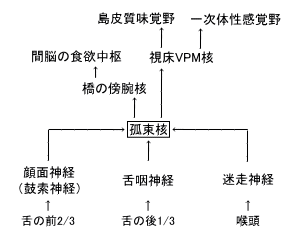
| 枴錛偺庬椶 | 巁枴 | 嬯枴 | 墫枴 | 娒枴 |
| 愩偱偺嬊嵼 | 懁曽 | 屻曽 | 慜曽 | |
| 枴妎傪姶偠傞巇慻傒 | 愭抂枌偺揹埑僎乕僩K+僠儍僱儖偺H+偵傛傞幷抐仺枴錛嵶朎偑扙暘嬌仺掙晹偐傜恄宱揱払暔幙傪曻弌 | 愭抂枌偺G僞儞僷僋偺妶惈壔仺儂僗儂僕僄僗僥儔乕僛妶惈壔仺cAMP伀仺掙晹偺Ca2+僠儍僱儖傪奐偔仺枴錛嵶朎偑扙暘嬌仺掙晹偐傜恄宱揱払暔幙傪曻弌 | 愭抂枌偐傜偺Na+棳擖偵傛傞扙暘嬌仺掙晹偐傜恄宱揱払暔幙傪曻弌 | 愭抂枌偺G僞儞僷僋偺妶惈壔仺cAMP擹搙仾仺僞儞僷僋僉僫乕僛A偺妶惈壔仺掙晹偺K+僠儍僱儖偺晄妶壔仺枴錛嵶朎偑扙暘嬌仺掙晹偐傜恄宱揱払暔幙傪曻弌 |