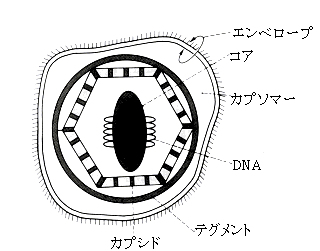
| �E�C���X�̓��� |
| �E�C���X���q�̊�{�\���ƌ`�� |
| �E�C���X�̕��� |
| dsDNA�E�C���X�AssDNA�E�C���X�AdsDNA(RT)�E�C���X�AdsRNA�E�C���X� ss(�|)RNA�E�C���X�Ass(�{)RNA�E�C���X�Ass(�{)RNA(RT)�E�C���X |
| �j�_ | �� | �E�C���X�� | ���� | ���� |
| dsDNA | �|�b�N�X�E�C���X | �V�R���i1980�N�ɍ���錾���Ȃ��ꂽ�j | �@ | |
| �w���y�X �E�C���X | �P���w���y�X�E�C���X �P�^�iHSV-1�j | ���O�w���y�X�A����w���y�X | �Q�m���͐���Q�{��DNA�B�Q�m���̒���100����ȏ�̑傫�����������z��������̂����� | |
| �P���w���y�X�E�C���X �Q�^�iHSV-2�j | ����w���y�X | |||
| �����я��v�]�E�C���X �iVZV�j | �����A�я��v�] | |||
| �q�g�T�C�g���K���E�C���X �iHCMV�j | ��V�����זE�����̏ǁA �A����P�j�� | |||
| EB�E�C���X�iEBV�j | �`�����P�j�ǁA�o�[�L�b�g �����p��A������� | |||
| �A�f�m�E�C���X | �@ | ��ᇃE�C���X | ||
| �p�|�o �E�C���X | �p�s���[�}�E�C���X | �@ | ��DNA�̎�ᇃE�C���X | |
| ssDNA | �p���{�E�C���X | �@ | �@ | |
| dsDNA (RT) | �y�p�h�i �E�C���X | �a�^�̉��E�C���X�iHBV�j | �}���a�^�̉��A�����̉��A �̍d�ρA�������̊� | �@ |
| �j�_ | �� | �E�C���X�� | ���� | ���� | |
| dsRNA | ���I �E�C���X | ���^�E�C���X | �@ | 11���߂̂Q�{��RNA������ | |
| ss(-)RNA | �p���~�N�\ �E�C���X | ���]�E�C���X | �E�C���X���ǁA�������A�x���A���]��]�� | ���j���זE���`������ | |
| �I���g�~�N�\ �E�C���X | �C���t�� �G���U �E�C���X | �`�^ | �C���t���G���U | ��`�q���W���߂ɕ�����Ă��� | |
| �a�^ | |||||
| �b�^ | �V���� | ||||
| ss(+)RNA | �s�R���i
�E�C���X | �|���I�E�C���X | �@ | �@ | |
| �`�^�̉��E�C���X �iHAV�j | �����̕s���������A����҂ł̏d�lj� | �o�������ɂ�芴�� | |||
| �t���r �E�C���X | �b�^�̉��E�C���X �iHCV�j | �����̉��A�̍d�ρA�������̊� | �\�h�E���ÂƂ���� | ||
| ss(+)RNA (RT) | ���g�� �E�C���X | HTLV-1 | ���l�s�זE�����a�iATL�j | �Q�m�����זE�̐��F��DNA�̒��ɑg�ݍ��� | |
| �q�g�Ɖu�s�S �E�C���X�iHIV�j | AIDS | ||||
| �E�C���X�̑��B |
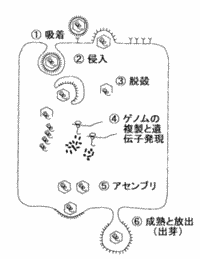
| �@�z���F | �h��זE��̃��Z�v�^�[�ƌ�������B |
| �A�N���F | (�@)�����̃E�C���X�̓G���h�T�C�g�[�V�X�ɂ���āA(�A)�G���x���[�v�����E�C���X�̓G���x���[�v�ƍזE����Z�������āA(�B)���^�E�C���X�Ȃǂ͒��ڍזE���f���āA�זE���ɓ���B |
| �B�E�k�F | �J�v�V�h��������E�C���X�Q�m�����זE�����ɕ��o�����B |
| �C�Q�m���̕����ƈ�`�q�����i�^���p�N�����j�F �������̃O���[�v�ɕ������B | |
| (�@)�N���X�T�F | ���N�V�j�A�E�C���X��A�f�m�E�C���X�̂悤�ɁA�Q�{��DNA���Q�m���Ƃ���E�C���X�BDNA�����ƈ�`�q�����̗l���͐^�j�����Ɠ����B |
|
���� �Q�{��DNA �\�\�� mRNA �\�\�� �^���p�N�� ���\�\�\�� |
| (�A)�N���X�U�F | �p���{�E�C���X�ȂǂP�{��DNA���Q�m���Ƃ���E�C���X�B�Q�m��DNA�͑����̏ꍇ�A(�|)���ł���B�����ɂ͏h��זE��DNA�|�������[�[��p���A�Q�{��DNA���o�čĂ�(�|)��DNA�ƂȂ�B��`�q�����́A�Q�{��DNA����h��זE��RNA�|�������[�[�𗘗p���āAmRNA���������邱�Ƃɂ��s����B |
|
���� (�|)��DNA �\�\�� �Q�{��DNA �\�\�� mRNA �\�\�� �^���p�N�� ���\�\�\�\�\�\�\�\�\�\�\�� |
| (�B)�N���X�V�F | ���I�E�C���X�ȂǁA���ߏ�Q�{��RNA���Q�m���Ƃ���E�C���X�B�Q�m����(�|)���𒒌^�ɃE�C���X��RNA�|�������[�[�ɂ�葽����(�{)��RNA����������A���ꂪmRNA�Ƃ��Ĉ�`�q������S���B����A���ꂪ(�|)��RNA�����̒��^�ƂȂ�A�E�C���X��RNA�|�������[�[�ɂ��Q�{��RNA�ƂȂ�B |
|
���� �Q�{��RNA �\�\�� mRNA �\�\�� �^���p�N�� ���\�\�\�\�\�\�\�\�\�\�� |
| (�C)�N���X�W�F | �s�R���i�E�C���X�A�g�K�E�C���X�A�t���r�E�C���X�ȂǁA(�{)��RNA���Q�m���Ƃ���E�C���X�B�Q�m��RNA�͂��̂܂�mRNA�Ƃ��ċ@�\����B�Q�m��RNA�̕�����(�|)��RNA�̒��ԑ̂𒒌^�Ƃ��čs���B |
|
���� (�|)��RNA �\�\�� (�{)��RNA �� mRNA �\�\�� �^���p�N�� ���\�\�\�\�\�\�\�\�\�\�\�� |
| (�D)�N���X�X�F | �I���g�~�N�\�E�C���X�A�p���~�N�\�E�C���X�Ȃ�(�|)��RNA���Q�m���Ƃ���E�C���X�B�E�C���X��RNA�|�������[�[�ɂ��(�{)��RNA����������AmRNA�Ƃ��ċ@�\�������A�Q�m������(�{)��RNA�𒒌^�ɃQ�m��RNA�����������B |
|
���� (�|)��RNA �\�\�� mRNA �\�\�� �^���p�N�� ���\�\�\�\�\�\�\�\�\�\�� |
| (�E)�N���X�Y�F | (�{)��RNA���Q�m���Ƃ��A�t�]�ʂɂ��DNA���������A�h��DNA�ɑg�ݍ��܂��i�v���E�C���X�j���g���E�C���X�B |
|
�@�@�@�@�@�@�@�@�@���\�\���^���p�N�� ���� (�{)��RNA �� mRNA �\�\�� (�|)��DNA �\�\�� �Q�{��DNA ���\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�� |
| (�F)�N���X�Z�F | �����I�ɂP�{���̕���������Q�{��DNA���Q�m���Ƃ���w�p�h�i�E�C���X�B�Q�m���̕����ɂ́A�Q�m����菭������(�{)���i�v���Q�m���j�̒��ԑ̂�K�v�Ƃ��A�t�]�ʂɂ��DNA�����������B����A��`�q�����́A�Q�m��DNA�����S�ȂQ�{���ɏC�����ꂽ��A�h��זE��RNA�|�������[�[�ɂ��mRNA����������A��`�q�������s����B |
|
���� (�|)��DNA �\�\�� �Q�{��DNA �\�\�� mRNA �\�\�� �^���p�N�� ���\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�\�� |
| �D�A�Z���u���F | �������ꂽ�Q�m����^���p�N�Ȃǂ��g�ݗ��Ă��A�E�C���X���q���`�������B���ꂪ�N����ꏊ�i�j�����זE���j�̓Q�m���̕����ꏊ�ɂ���ĕς��B |
| �E���n�F | �V���ɎY�����ꂽ�E�C���X�������\�����悤�Ɋ������Ă����ߒ��B |
| �F���o�F | �זE�O�ɏo��B |
| ��ᇃE�C���X |
| �E�C���X�̓`�d�o�H�Ɗ����l�� |
| SEO | [PR] ����!�����u���O �����z�[���y�[�W�J�� �������C�u���� | ||
